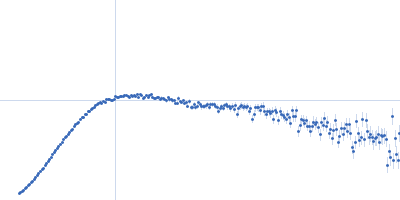
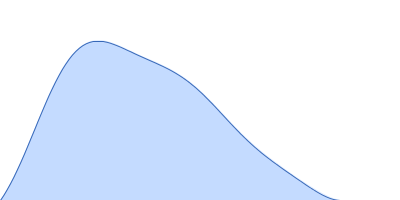
Synchrotron SAXS data from solutions of Truncated P5abc subdomain from tetrahymena ribozyme: Static 0.50mM MgCl2 in 0.5mM MgCl2 20mM KCl 10mM KMOPS 20uM EDTA, pH 7 were collected on the G1 beam line at the Cornell High Energy Synchrotron Source (CHESS; Ithaca, NY, USA) using a CCD Finger Lakes CCD detector at a sample-detector distance of 1.5 m and at a wavelength of λ = 0.109 nm (I(s) vs s, where s = 4πsinθ/λ and 2θ is the scattering angle). Solute concentrations ranging between 0.8 and 1.5 mg/ml were measured at 20°C. 40 successive 10 second frames were collected. The data were normalized to the intensity of the transmitted beam and radially averaged; the scattering of the solvent-blank was subtracted and the different curves were scaled for protein concentration. The low angle data collected at lower concentrations were extrapolated to infinite dilution and merged with the higher concentration data to yield the final composite scattering curve.
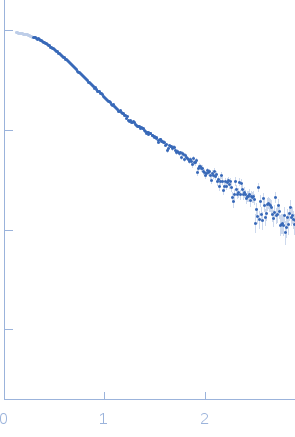 s, nm-1
s, nm-1